


樹状細胞(dendritic cell, DC)による免疫系の制御
生体防御とは、一義的にはウイルス細菌などの病原性の微生物等の侵襲を防ぐと共にそれらを排除することである。
この過程には、初期に誘導される非特異的な自然免疫応答とそれに続いて誘導される特異的な獲得免疫応答が作用している。
自然免疫応答には微生物侵襲部位の体細胞はもとより、骨髄肝細胞に由来する好中球、マクロファージ、ナチュラルキラー細胞(NK細胞)、NKT細胞など多くの細胞が関与している。一方、獲得免疫応答はT細胞やB細胞などのリンパ球細胞に担われる応答であるが、これらの細胞が機能するためには抗原情報を提示する細胞が必要とされる。
この役割を担う細胞は、抗原提示細胞と称され、マクロファージ、B細胞、樹状細胞がプロフェッショナル抗原提示細胞として扱われてきた。
しかし、ナイーブなT細胞を刺激することが出来るのは樹状細胞のみである。
樹状細胞は生体に広く存在し、それぞれの末梢組織で外来性の異物だけでなく自己成分を取り込み所属リンパ節に移動する。
この過程で取り込んだ物質を分解し主要組織適合抗原(MHC)上に提示することにより特異的T細胞を活性化し、種々の免疫応答を惹起する。
ところが時として非感染性の外来物質や自己成分に対しても応答し、アレルギーや自己免疫疾患などを引き起こすことも知られている。
これまでの研究から、樹状細胞集団は複数のサブセットより構成され、その成熟段階や分化経路が異なることが明らかになっている。
しかもこれらは異なる機能や活性をもち、免疫応答に対してそれを増強するだけでなく、抑制する作用を及ぼすことも知られるようになっている。
生体応答学分野では、このような樹状細胞に焦点を当て、自然および獲得免疫応答成立過程における “免疫の制御細胞”としての役割を解明するための研究を進めている。
これにより自己免疫疾患および慢性の免疫関連疾患に関する新たな知見が得られるものと期待される。
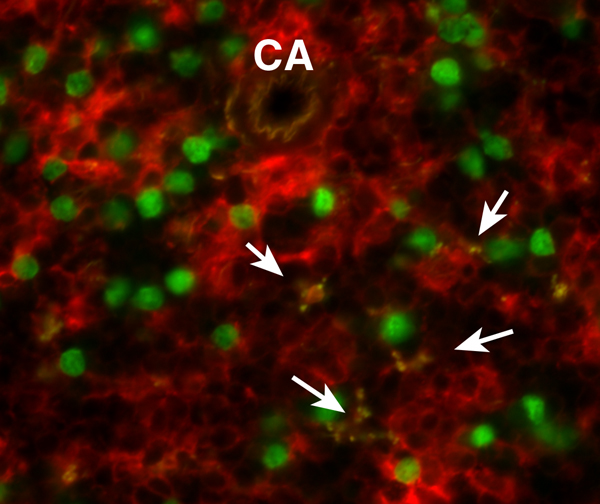
Fig.1. 脾臓DCによる死細胞の取り込み
生体内樹状細胞による死細胞の取り込み
様々な粒子状抗原を取り込む能力はマクロファージに見られる機能と考えられてきた。
しかし、樹状細胞も種々の物質を多様な機構で取り込む機能を持っている。
一般に、マクロファージはそれぞれの組織で移動することなく、そこに生じた物質を捕食し消化分解して生体にとって無害なものとする。
これに対して、樹状細胞は末梢組織において抗原を取り込んだ後に、リンパ器官へと遊走し、分解産物をT細胞へと提示する。
この過程で、有害な抗原や病原体に対しては免疫応答を誘導するが、自己成分や無害な物質に対しては免疫寛容を誘導することができる。
生体組織や器官では、絶えずアポトーシスによる細胞死が見られ、これらを捕捉して末梢における組織特異的免疫寛容を誘導することによって生体の恒常性を維持することも樹状細胞の担う重要な機能である。
生体内の樹状細胞は細胞表面に発現している分子群の違いによって複数のサブセットに分けられるが、マウスでは特に、CD8α、CD11b、CD205の発現を指標として大きく2つのDCサブセットの存在が知られている。
これらは共に可溶性抗原やラテックス粒子を等しく取り込むことが出来るが、リンパ系器官において死細胞はCD8α+、CD11blow、CD205+の樹状細胞のみが貪食して入ることが示され、そのため死細胞に由来する抗原はこのサブセットの樹状細胞によってのみ提示されることも明らかになった。これをもとに、樹状細胞サブセットにおける死細胞認識に働く受容体の検討を行っている。
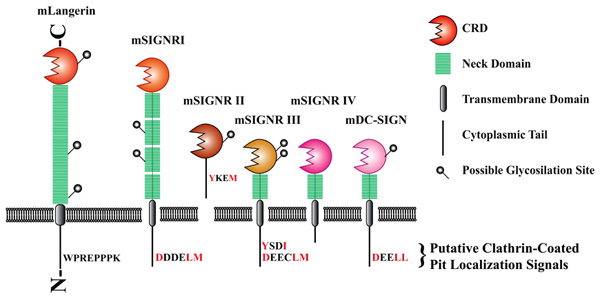
Fig.2. 現在解析中のC型レクチン
微生物認識および取り込みに働くレセプターに関する研究
末梢組織に分布する樹状細胞は、免疫監視細胞として細菌やウイルスなど侵襲異物を認識し、それらを排除すべく働いている。
そのために、種々のレセプターを発現し、それらを認識・捕捉すると同時に活性化され、自然免疫応答ならびに獲得免疫応答の増強に働いている。
微生物等を認識する受容体はパターン認識分子と称され、TLRs(Toll-like receptors)やC型レクチンが知られる。
このうち、C型レクチンは病原体上の糖のパターンを認識し見分けるだけでなく、自己の糖タンパク質と結合することで樹状細胞の分化や移動にも関与している。
先に我々は、表皮ランゲルハンス細胞に発現するマウスランゲリン(CD207)、およびヒトDC-SIGN(CD209)のマウスホモログ(計5種)をクローニングした(Fig. 2)。
現在、これらのC型レクチンにより認識されるリガンド、およびこれらを感染のターゲットに利用する病原微生物の検索を進めている。また、HIV等レクチンを介して免疫系から“escape”する術を身につけた病原体が知られているが、この場合レクチンを介して伝わる細胞内シグナルが他の病原体の場合と異なるのではないかと考え、この点についても研究を進めている。
抗原提示とT細胞の活性化
抗原特異的な免疫応答の成立には、T細胞がその抗原受容体を介して抗原提示細胞上に発現されたMHC分子に結合した抗原ペプチドを認識しなければならない。
しかし、これら両分子間の作用のみでT細胞が活性化されるのではなく、細胞接着分子や共刺激分子(co-stimulatory molecules)の作用によって、この認識がより強固なものとされる。
これらの分子間相互作用は分子間クラスターという特殊な場の中で成立する。
このような場を免疫シナプスと呼び、細胞膜上に形成されるコレステロールやスフィンゴ脂質に富むマイクロドメインがその場となる。
しかしながら、免疫シナプスというマクロドメインの形成がMHCおよび共刺激分子/接着因子を高発現しかつ特有な機能を有する樹状細胞の機能において、どのような役割あるいは作用をしているのか、また樹状細胞の成熟活性化とどのように関連しているのかは不明な点が多い。
そこで、異なった分化段階の骨髄由来樹状細胞を用いて、細胞表面上のMHCおよび共刺激因子を含むマイクロドメインの形成を検討し、これが効果的な免疫シナプスの形成に関与している可能性を検討している。
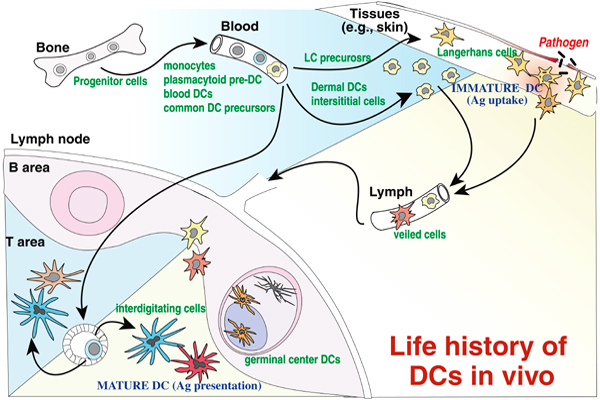
Fig.3. DCサブセットの生活史
生体内における樹状細胞サブセットの解析
生体内には表現型や機能から識別可能ないくつかの樹状細胞サブセットが存在している。
しかもそれらは定常状態においても常に骨髄幹細胞より供給され、所属リンパ節に移動している(Fig. 3)。
しかもその過程で捕捉した種々の抗原をリンパ組織においてに提示している。
定常状態においてはこのような抗原提示は、末梢免疫寛容の誘導と維持に重要役割を担うことが我々の研究で明らかになってきている。
しかしながら、いかなる樹状細胞サブセットが免疫寛容に関わるのか、また特異的T細胞の免疫不応答状態はどのような機構によって成立しているのかなど不明な点が多い。
樹状細胞サブセットの機能を考慮しつつ、特異的T細胞の除去、不活化さらには制御性/抑制性T細胞の誘導などの可能性の検討を行っている。
一方、定常状態はもとより炎症性応答が誘導された場合の樹状細胞の移動に関する研究は進んでおり種々のケモカインの役割も明らかになってきている。
しかし、個体全体における動態や寿命さらにそれらを制御する要因に関しては情報が少ない。
この点を考慮し、生体内各器官に分布する樹状細胞サブセットの構成比と生存期間、さらにはそれらの特異的あるいは非特異的免疫制御細胞としての機能を検討している。